Critique of the Scientific Evidence for Fisheries Benefits of MRs
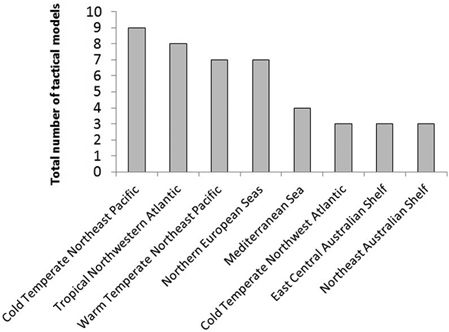
Fig. 5.1
Classification scheme for the empirical and theoretical MPA biological literature
5.3 Results
5.3.1 Empirical Studies
Number of Empirical Studies
As literature on MPAs has continued to expand exponentially, the proportion of empirical studies has increased relative to both theoretical and review/note type literature (see Fig. 5.2). This was observed for all empirical field studies that have been undertaken in MPAs (n = 448) and for those empirical studies that have examined the effect of protection only (n = 310) (see Fig. 5.3). Recently, however, the annual publication rate of theoretical papers has been increasing at a similar rate to that of empirical studies, due to a leap in the publication of modelling studies.
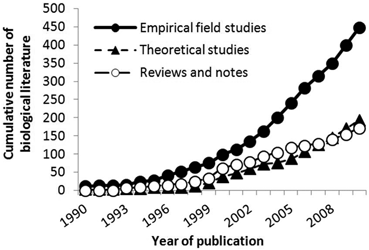
Fig. 5.2
Publications concerned with the biology of MPAs in the published literature, 1990–2010: comparison of the number of empirical, theory, and reviews/notes studies. To aid visualisation, papers 1977–1989 (n = 8) were categorised as 1990
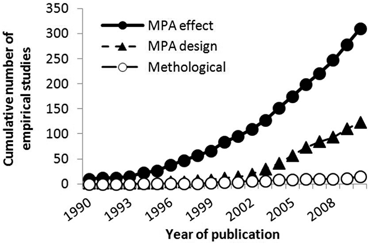
Fig. 5.3
Empirical studies broken down by type: whether they investigated the effect of protection (MPA effect); gained evidence on the movement/distribution of species/habitats to inform MPA design (MPA design); or investigated a methodological problem
Type of MPA Studied
Eighty-seven percent of the empirical literature has focused on effects of MRs (see Fig. 5.4). The remaining 13 % of studies have mainly focused on effects of no-trawl areas predominantly established over soft-bottom habitats in temperate seas (e.g. Murawski et al. 2000; Jaworski et al. 2010), or MPAs that only allow recreational users (Denny and Babcock 2004; Shears et al. 2006). Twenty-five percent of the empirical literature has come from the 10 MRs shown in Fig. 5.5. As of 2010, around 170 MRs had been studied, 30 of which were located in the Mediterranean Sea.
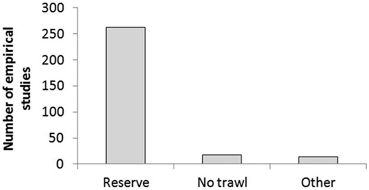
Fig. 5.4
Type of MPA studied: ‘Reserve’ defined as an area where no fishing occurred; ‘No trawl’ as an area where towed ground gear was prohibited; and ‘Other’ defined as areas that only allowed recreational users
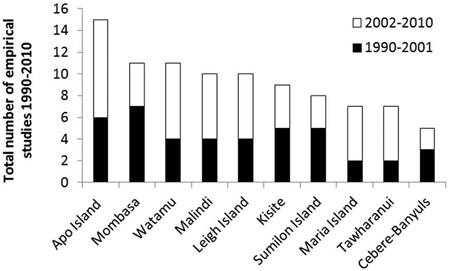
Fig. 5.5
Top 10 MPAs studied 1990–2010. Note that all are MRs
Ecosystems and Habitats Studied
When the empirical literature is broken down by ecosystem type, more studies have been undertaken in the tropics (n = 119) and warm temperate ecosystems (n = 116), and more than half of this research effort has been undertaken in 25 MRs in the Mediterranean Sea. The publication rate of cold temperate ecosystems research lags well behind that of tropical and warm temperate ecosystems (Fig. 5.6). Reef-type habitats have been most studied (see Fig. 5.7a, n = 228) with only 16 % of studies being carried out over soft habitats (see Fig. 5.7b, n = 43 including no-trawl areas). There has been roughly the same research effort applied over soft habitats in warm temperate and cold temperate ecosystems, though this is mainly due to the study of the effects of large-scale groundfish closures that are predominantly located over soft ground (see Fig. 5.7b).
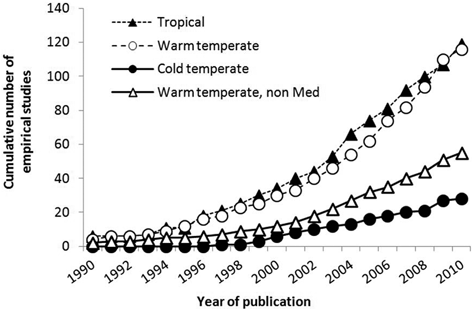
Fig. 5.6
Number of empirical field studies undertaken in MRs only by ecosystem type. The subset of warm-temperate studies conducted outside the western Mediterranean Sea is plotted separately
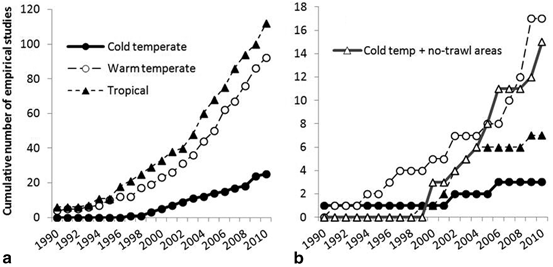
Fig. 5.7
Number of empirical field studies that have measured the effect of an MR over (a) hard (reef) habitats, and (b) soft habitats (with studies that have examined cold temperate no-trawl areas also shown)
Main Groups of Species Studied
Reflecting the type of habitat surveyed, the main group of species studied were coral and rocky reef fish communities (see Fig. 5.8), which comprised 45 % of the focus species of all empirical studies. Reef crustaceans (n = 22), coral reef fish predators (n = 22) and molluscs (n = 19) have been the subject of 20 % of empirical studies, while temperate soft-bottom fish communities have been the subject of only 5 % of empirical studies. No MPA effect studies were found for charismatic marine megafauna, although a few studies relevant to the design of MPAs did have empirical data showing the distribution and movement of turtles (de Segura et al. 2003), birds (Louzao et al. 2006; Terauds et al. 2006), and cetaceans (Canadas et al. 2005; Slooten et al. 2006) in MPAs and their surrounds.
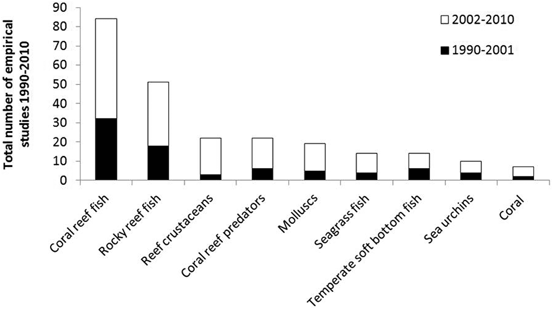
Fig. 5.8
Main focus organism(s) of MPA effect studies
Empirical Research Effort per Marine Province
Four marine provinces stand out in terms of the number of empirical studies that have been published on MPAs (see Fig. 5.9): the Mediterranean Sea (n = 70); the Tropical Northwest Atlantic (n = 33); the Western Indian Ocean (n = 24); and the Western Coral Triangle (n = 23). Cold temperate marine provinces that have been moderately studied (between 6 and 11 publications) include Northern European Seas, the cold temperate Northwest Atlantic, and the cold temperate Northeast Pacific. Around half of the marine provinces have not been studied at all, and there are notable information gaps for Asia, Western Africa and the Southern Ocean.
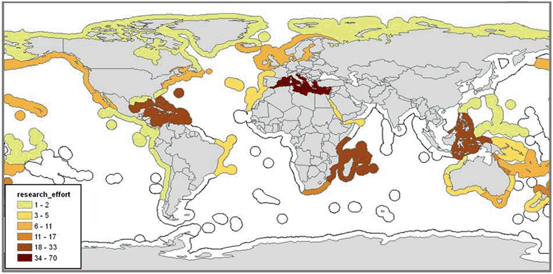
Fig. 5.9
Research effort (defined as number of empirical studies) per marine province: marine provinces with no colour have no MPA effect studies
‘Quality’ of Empirical Field Studies
Fifty four percent of empirical MR studies only presented a snap shot, having measured the effect of protection at a single point (i.e. a season) in time. Of these snap-shot studies, 25 % used only one fished control area to attribute a difference between sites to an effect of protection, and less than half of these explicitly tried to take into account effects of habitat in their survey design. Forty six percent of studies took inter-annual variation into account in their design (see Fig. 5.10), but only 12 % measured effects of protection over time periods > 10 years.
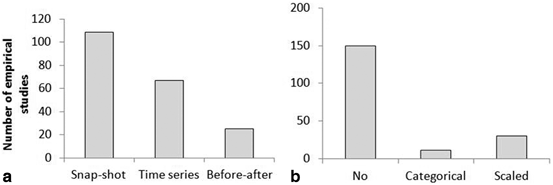
Fig. 5.10
Temporal aspects of empirical literature investigating MR effects (a) and number of studies that quantified habitat and used this as a covariate in their analysis (b)
Twelve percent of studies had before–after data (BACI); the design and focus of these studies are summarised in Table 5.1. The majority of before-after studies had measured effects of protection on abundance and biomass of coral reef fish species, though only one before-after study measured the effect of spillover (Francini-Filho and Moura 2008). Habitat was categorised (n = 11) or scaled (n = 30) by only 21 % of empirical studies.
Table 5.1
Summary of before-after studies of marine reserves, and number of temporal replicates per study (minimum two)
Reference(s) | Location | No. of reserves | No. of temporal replicates | Study focus |
|---|---|---|---|---|
Russ and Alcala (1989, 1998, 2003, 2004) | Sumilon and Apo Islands, Philippines | 2 | 2, 5, 13 | Coral reef fish species richness, density and biomass |
Bennett and Attwood (1991) | De Hoop, South Africa | 1 | 3 | CPUE [catch-per-unit-effort] of rocky reef fish assemblage |
Wantiez et al. (1997) | New Caledonia | 5 | 2 | Coral reef fish species richness, density and biomass |
Galal et al. (2002) | Nabq, South Sinai, Egypt | 5 | 3 | Density and size structure of commercially targeted grouper, emperor and snapper |
Nardi et al. (2004) | Houtman Abrolhos Islands, Western Australia | 4 | 6 | Density of coral trout and wrasse |
Claudet et al. (2006) | Couronne, France | 1 | 3 | Rocky reef fish species diversity and abundance |
Hawkins et al. (2006) | St Lucia, Caribbean | 4 | 7 | Commercial coral reef fish species biomass |
Lincoln-Smith et al. (2006) | Solomon Islands | 1 | 6 | Abundance and size of commercial coral reef invertebrate |
Francini-Filho and Moura (2008) | Eastern Brazil | 1 | 5 | Biomass, size, and spillover of coral reef fish |
Table 5.2 summarises the factors that determine the strength of an MPA effect and which should be taken into account in the design of a monitoring programme.
Table 5.2
Factors determining the strength of an MPA effect
Factor | Comments |
|---|---|
Habitat heterogeneity | MPAs may be located over habitats that are comparatively resource rich (Hilborn 2002) or resource poor (Edgar et al. 2009). An ecological effect could be attributed to protection when it is instead due to habitat differences. To counter this, studies should use BACI designs (Underwood 1993) to increase the strength of their inference |
Biological life histories | Individual species life-history traits strongly affect how they will respond to protection. Species may grow slowly so that any significant change will not be detected for years (Barrett et al. 2007). Alternatively, fish species may be highly mobile, so that an MPA has little or no protection effect (Shipp 2003). Survey designs and the amount of sampling effort needed to detect an effect will need to take species movement into account (Rotherham et al. 2007) |
Environmental change | Regional environmental change and its effect on fish growth and recruitment may confound the interpretation of the effect of a closed area (Holland 2000). Such a problem could only be overcome through long-term monitoring |
Past management history | The extent to which a fish stock has been exploited and habitat modified by fishing will influence the size of an ecological effect detected in an MPA relative to control locations and baseline at t = 0 |
Current management history | Displaced fishing effort outside the MPA may lead to a greater intensity of fishing in its surrounds and lead the researcher to detect a greater effect of protection due to the deterioration of fish stocks and habitat outside (Hilborn 2002). Illegal fishing may also reduce the size of ecological effects (Bloomfield et al. 2012) |
5.3.2 Theoretical Studies
In the theoretical (i.e. modelling) studies, there was a greater abundance of strategic (n = 130) than tactical models (n = 56). The publication of strategic models increased rapidly during 1999–2000, and it was not until 2008–2009 that the publication trajectory of tactical models started to match that of strategic models—roughly a 9-year lag (see Fig. 5.11). Seventy percent of the strategic models were not calibrated to specific species: for those that were, 13 % derived their parameters from tropical species (mainly coral reef fish), compared to 7 % from warm temperate species, and 8 % from cold temperate species (mainly cod).
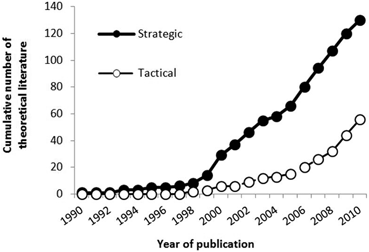
Fig. 5.11
Number of theoretical studies by model type
For temperate ecosystems, twice as many papers showed tactical models (n = 40) than showed strategic models (n = 20) (see Fig. 5.12), many of these informing the design of MPA networks along the northeast Pacific coast of the USA (Ban 2009; Ban and Vincent 2009; Airame et al. 2003), and predicting the effects of groundfish closures (Horwood et al. 1998; Holland 2000; see Fig. 5.13 and Table 5.3).
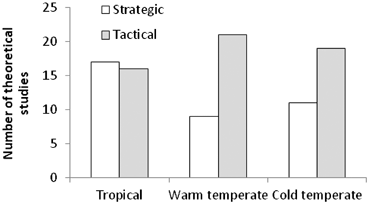
Fig. 5.12
Total numbers of theoretical studies by ecosystem. Note that 70 % of strategic models were completely abstract, whereas by their very nature all tactical models were developed for real-world problems

Fig. 5.13
Total numbers of tactical models per marine province
Table 5.3
Some examples of tactical models potentially used to inform local fisheries and biodiversity conservation policy
Reference | Area/MPA | Findings |
|---|---|---|
Horwood et al. 1998 | Trevose spawning grounds for sole, Celtic Sea | Found that the closure of a sole spawning ground may be ineffective if sole remain free to be caught elsewhere, and the catch of sole outside the closed area is still high. Closed areas will be useful in protecting aggregations of juvenile fish |
Holland 2000 | New England groundfish closures | Impacts of closures will vary across species, sometimes increasing yields for some and decreasing yields for others |
Airame et al. 2003 | California Channel Islands | Identified reserve network scenarios that would represent all habitats whilst minimising socio-economic costs to stakeholders |
Stewart et al. 2003 | South Australia | Ad hoc placement of marine reserves may compromise effective conservation of marine biodiversity |
Kjaersgaard and Frost 2008 | Plaice box, North Sea | Plaice box largely ineffective; need to reduce the fishing effort of smaller vessels still fishing in the closed area to achieve a profitable fishery with a biomass above the reference point Bpa |
5.4 Discussion
Four themes emerged from this literature review of MRs: drawbacks of targets; skewed focus of literature; mixed evidence; and difficulties of enforcement.
5.4.1 Drawbacks of Targets
Although few scientists explicitly claim that MRs are a panacea for all marine resource management problems (Roberts 1997; Lubchenco et al. 2003; Pitcher and Lam 2010; Mora 2011), there is no escaping the fact that MRs have dominated the literature on marine resource management during the last 20 years (see Chap. 1), and they have been advocated more frequently than other new management strategies such as marine spatial planning (MSP) or ecosystem-based management (EBM) (Halpern et al. 2010). Yet only 1.6 % of the ocean is designated as MPAs and only 0.2 % as MRs (Wood et al. 2008), which is a long way from meeting current international targets. In 2006 the CBD called for 10 % of the oceans to be designated as MPAs by 2010, but, according to Wood et al. (2008), at the (then) current rate of progress, this target would not be achieved until 2050. However, a more up-to-date assessment by Spalding et al. (2013, p. 231), drawing attention to ‘The dramatic acceleration of MPA coverage in recent years’, in particular the increase in MPA designations of vast areas of the ocean during the past 5 years (including South Orkney, South Georgia, Chagos, and Motu Motiro Hiva), and counting fisheries protected areas and other kinds of site protection, claimed that ‘Rates of progress suggest that it may well be possible to attain 10 per cent coverage by 2020’. Moreover, as Jones (2014, p. 8) notes, ‘only the IUCN target specifically requires no-take MPAs…The other targets, including under the CBD, could be met by partially protected MPAs that include provisions for some extractive activities’ (italics in original). And if areas are counted where temporary (e.g. seasonal) protection is afforded, and/or real time restrictions are imposed to protect spawning or juveniles, the global targets may well be feasible.
Nevertheless, there are drawbacks with targets. One drawback is that the targets may have little scientific justification—‘ecologically irrelevant’ (Metcalfe et al. 2013, p. 10)—because they have been virtually picked out of the air. One conservation scientist, who has complied IUCN guidelines, explained that the 10 % figure that was initially adopted by the CBD was formulated arbitrarily over dinner: a figure of 5 % was suggested but it was thought that 5 % would be more likely to be achieved if a 10 % target were set (pers. comm). On the call for a 20 % global target, Agardy et al. (2003, p. 361, 359, 360) stated that ‘The 20 % figure has been elevated to a dogmatic standard for a minimum proportion of a type of ecosystem that must be delineated as no-take MPA in order for the MPA to be effective in protecting natural resources’, but it was originally ‘extrapolated from very specific localized studies of particular fisheries within particular habitats—not from representative community ecology data from a wide variety of habitat types…Yet the 20 % figure has been adopted as the mantra of some MPA advocates targeting a wide range of objectives under a diverse spectrum of ecological and social conditions…Does the ease of convenience in selecting a single rule-of-thumb figure for all situations run the risk of selecting meaningless threshold targets?’. Similarly, Planes (2011, p. 327) reported that ‘In the late 1990s a magic number came out of some modelling studies suggesting that 30 % of the coastlines should be included in MPAs in order to ensure maintenance of the overall ecosystem functioning’. The US Coral Reef Task Force recommended that a minimum of 20 % of the southern Atlantic coast of the USA should be included in MRs on the basis that it would protect 20 % of the spawning biomass, a threshold below which stocks were likely to collapse (Bohnsack et al. 2000). Studies have claimed that setting aside 10–30 % of the sea as MRs in regions dominated by fishing impacts, can improve overall ‘ocean health’ by reducing the total cumulative impact on the ecosystem by 15–20 % (Halpern et al. 2010). But little scientific proof has been provided for these quantitative predictions. Moreover, ‘overall ecosystem functioning’, ‘ocean health’ and ‘total cumulative impact’ are highly contested concepts that are invariably left undefined. Wood et al. (2008, p. 349) claimed that targets are politically not ecologically driven: ‘Targets have historically been justified in terms of political expediency rather than ecological knowledge’ (see also De Santo 2013). Targets do not necessarily synchronise with areas requiring protection. As both De Santo (2013) and Metcalfe et al. (2013) noted, percentage area targets falsely imply that biodiversity is evenly distributed across the seas, and that the marine conservation needs of a country coincide with a particular percentage of protected sea area.
Another drawback of targets is that even where the targets do have scientific justification they may require enormous areas of MRs, which are politically untenable. Kaiser (2005) pointed out, for example, that a study had shown that even excluding fishers from 25 % of the North Sea would have only a negligible effect in protecting cod spawning stock biomass, while another study proposed that such highly mobile species required MRs spanning up to 65 % of particular seas.